The Freshwater Mussels (Unionoida) of the World (and other less consequential bivalves)
FM(U)otW(aolcb) is the web version of the MUSSEL Project Database. Follow the links to browse the data or use the custom Google search field. Either way, you win!
Search the database pages:
Bivalvia | Unionida | Hyrioidea | Hyriidae | Hyriinae | Castaliini genus Castalia Lamarck, 1819: cladomics
13 Recent valid species (13 total species) Cladomics Summarynumber of species = 13 (13 total)
analyzed in 39 cladograms in 25 publications
maximum number of terminals in an analysis =
3 (mean = 1.15, median = 1) Basic Monophylynumber of cladograms that test basic monophyly
(i.e., more than 1 included species) = 5
maximum number of included terminals = 3, (mean =
2.2 median = 2)
proportion of cladograms that support basic monophyly
= 0.4
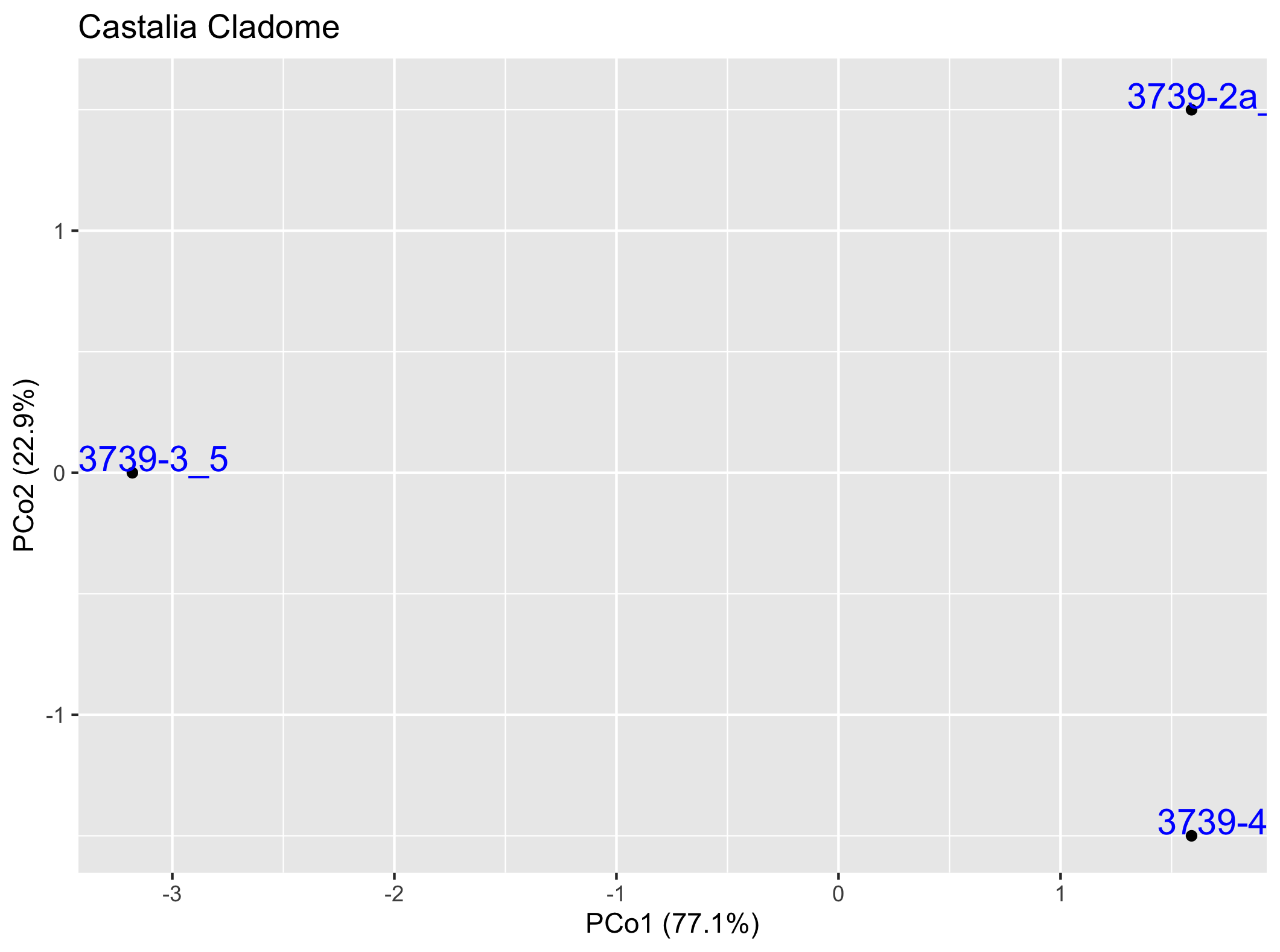
Pairwise Cladogram ClusteringThe graphs below depict in two dimensions all pair-wise ingroup distances within the cladome. See the Cladmoics FARQ for more information.
Campbell & Lydeard, 2026 [Publ 4810], fig. 6-14 ** Top Monophyly Cladogram ** ** Top Sister-Group Cladogram ** | iT | tT | bi | riS | oT | roT | roS | iD | | 3 | 5 | 6 | 6 | 629 | 2 | 2 | 0 |
+----------------- Castalia_ambigua__4 **
|
--+ +-------------- Callonaia
| |
+--+ +----------- Castalia_stevensi **
| |
+--+ +-------- Castalia_ambigua__3 **
| |
+--+ +-- Castalia_ambigua__1 **
| +--+
| | +-- Castaliella
+--+
| +-- Castalia_ambigua__2 **
+--+
+-- Castalia **
Santos-Neto et al., 2018 [Publ 3888], fig. 5 | iT | tT | bi | riS | oT | roT | roS | iD | | 2 | 2 | 0 | 2 | 3 | 1 | 1 | -- |
+----- Callonaia
--+
| +-- Castalia_ambigua **
+--+
+-- Castalia_stevensi **
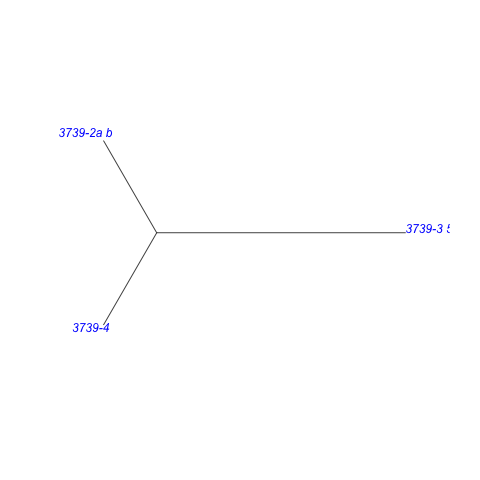
Santos-Neto et al., 2016 [Publ 3739], fig. 2a_b | iT | tT | bi | riS | oT | roT | roS | iD | | 2 | 4 | 0 | 5 | 18 | 2 | 2 | 4 |
+-------------- Castalia_stevensi **
|
--+ +-- Callonaia
| +--------+
| | +-- Castalia_ambigua__1 **
+--+
| +-------- Castalia_ambigua__4 **
+--+
| +----- Castalia_ambigua__2 **
+--+
| +-- Castaliella
+--+
+-- Castalia_ambigua__3 **
Santos-Neto et al., 2016 [Publ 3739], fig. 4 | iT | tT | bi | riS | oT | roT | roS | iD | | 2 | 4 | 0 | 5 | 18 | 2 | 2 | 2 |
+----------------- Castalia_ambigua__4 **
--+
| +-------------- Callonaia
+--+
| +----------- Castalia_ambigua__3 **
+--+
| +-------- Castalia_stevensi **
+--+
| +----- Castalia_ambigua__2 **
+--+
| +-- Castaliella
+--+
+-- Castalia_ambigua__1 **
Olivera-Hyde et al., 2020 [Publ 4249], fig. 10 | iT | tT | bi | riS | oT | roT | roS | iD | | 2 | 2 | 0 | 2 | 1 | 1 | 1 | -- |
+----- Callonaia
--+
| +-- Castalia_ambigua **
+--+
+-- Castalia_stevensi **
Bolotov et al., 2017 [Publ 3761], fig. S1 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 138 | 0 | 0 | -- |
Graf et al., 2015 [Publ 3619], fig. 1 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 19 | 0 | 0 | -- |
Hoeh et al., 2009 [Publ 2716], fig. 3 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 46 | 0 | 0 | -- |
Graf, 2002 [Publ 2651], fig. 2 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 17 | 0 | 0 | -- |
Roe & Hoeh, 2003 [Publ 2655], fig. 4f3 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 30 | 0 | 0 | -- |
Walker et al., 2006 [Publ 2666], fig. 6 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 35 | 0 | 0 | -- |
Hoeh et al., 2002 [Publ 2653], fig. 2 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 30 | 0 | 0 | -- |
Santos-Neto et al., 2016 [Publ 3739], fig. 3_5 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 2 | 0 | 2 | 6 | 2 | 2 | -- |
+-------- Callonaia
--+
| +----- Castalia_ambigua__2 **
+--+
| +-- Castalia_ambigua__1 **
+--+
+-- Castaliella
Hoeh et al., 2009 [Publ 2716], fig. 4 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 45 | 0 | 0 | -- |
Whelan et al., 2011 [Publ 3380], fig. 1-ML | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 35 | 0 | 0 | -- |
Huang et al., 2019 [Publ 3843], fig. S3 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 198 | 0 | 0 | -- |
Bogan & Hoeh, 2000 [Publ 2650], fig. 1 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 33 | 0 | 0 | -- |
Huang et al., 2019 [Publ 3843], fig. 2 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 200 | 0 | 0 | -- |
Whelan et al., 2011 [Publ 3380], fig. 1-MP | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 35 | 0 | 0 | -- |
Graf & Ó Foighil, 2000 [Publ 2649], fig. 1_2 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 10 | 0 | 0 | -- |
Graf & Cummings, 2006 [Publ 1968], fig. 2 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 46 | 0 | 0 | -- |
Bolotov et al., 2016 [Publ 3743], fig. 4 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 29 | 0 | 0 | -- |
Baker et al., 2003 [Publ 2690], fig. 5 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 7 | 0 | 0 | -- |
Hoeh et al., 2001 [Publ 2652], fig. 14f4 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 30 | 0 | 0 | -- |
Hoeh et al., 2009 [Publ 2716], fig. 5 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 45 | 0 | 0 | -- |
Campbell & Lydeard, 2026 [Publ 4810], fig. 3_5 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 139 | 0 | 0 | -- |
Wu et al., 2021 [Publ 4366], fig. 6 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 90 | 0 | 0 | -- |
Konopleva et al., 2017 [Publ 3807], fig. 6 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 30 | 0 | 0 | -- |
Baker et al., 2004 [Publ 2685], fig. 2-3 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 8 | 0 | 0 | -- |
Wu et al., 2021 [Publ 4366], fig. 2_I | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 90 | 0 | 0 | -- |
Guerra et al., 2019 [Publ 4168], fig. 1 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 40 | 0 | 0 | -- |
Huang et al., 2019 [Publ 3843], fig. S4 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 200 | 0 | 0 | -- |
Roe & Hoeh, 2003 [Publ 2655], fig. 4f2 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 30 | 0 | 0 | -- |
Bolotov et al., 2017 [Publ 3761], fig. S2 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 138 | 0 | 0 | -- |
Graf, 2000 [Publ 2647], fig. 1 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 16 | 0 | 0 | -- |
Pfeiffer & Graf, 2015 [Publ 3620], fig. 2 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 39 | 0 | 0 | -- |
Bolotov et al., 2016 [Publ 3743], fig. 2 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 29 | 0 | 0 | -- |
Hoeh et al., 2001 [Publ 2652], fig. 14f2 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 30 | 0 | 0 | -- |
Guerra et al., 2019 [Publ 4168], fig. 2 | iT | tT | bi | riS | oT | roT | roS | iD | | 1 | 1 | 0 | 1 | 40 | 0 | 0 | -- |
|